Ligação Gênica e Permutações
1. O que é ligação gênica?
Dois ou mais pares de genes alelos localizados em diferentes pares de cromossomos homólogos segregam-se independentemente. Portanto, esta é a condição de validade da segunda lei de Mendel.
Quando dois ou mais pares de genes alelos estão localizados em um mesmo par de cromossomos homólogos, eles não obedecem à lei da segregação independente. Afinal, durante a meiose irá haver uma tendência de que esses genes permaneçam unidos, quando o par de homólogos se separar, como mostra a figura abaixo.

Quando dois ou mais pares de genes alelos se localizam em um mesmo par de cromossomos, dizemos que eles apresentam ligação gênica (ou ligação fatorial). Os autores de língua inglesa dão a essa situação o nome de linkage. No entanto, há um fenômeno capaz de alterar essa tendência de união. É a permutação gênica (ou crossing-over), troca de fragmentos entre cromossomos homólogos, que pode acontecer na prófase da primeira divisão da meiose.
Quando dois pares de genes alelos estão situados de tal forma, em um par de homólogos, que não ocorre permutação entre eles, diz-se que há linkage total entre eles. Caso haja permutação, o linkage é parcial.
Em um caso de ligação gênica, não basta se conhecer o genótipo de um indivíduo. É necessário que se determine a posição relativa dos genes no par de homólogos. Por que isso é tão importante? Observe as duas situações mostradas a seguir:

Podemos notar que, embora as duas células possuam os mesmos genes, a sua posição, no par de cromossomos homólogos não é a mesma, o que determina a produção de tipos diferentes de gametas, na meiose.
Existem diversas formas de se indicar a posição dos genes no par de homólogos. As mais comuns são:

Uma outra forma de se indicar essa posição relativa dos genes é uma nomenclatura habitualmente usada pela química orgânica. O duplo-heterozigoto que tem os dois genes dominantes no mesmo cromossomo e os dois recessivos no outro (AB/ab) é chamado de heterozigoto "cis". O duplo-heterozigoto cujos genes dominantes estão em cromossomos diferentes do par de homólogos (Ab/aB) é o heterozigoto "trans".
2. Permutação ou crossing-over:
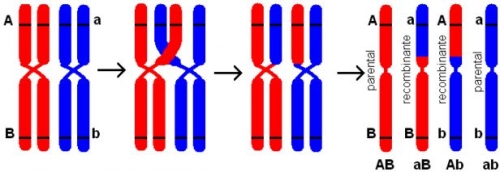
Teoricamente, os genes presentes em um mesmo cromossomo deveriam permanecer ligados uns aos outros, mesmo após a meiose. Porém, nem sempre isto ocorre. Muitas vezes é possível que os cromossomos troquem partes entre si durante a prófase da primeira fase da meiose.
Esse processo chamado de crossing-over ou permuta, ocorre quando os cromossomos homólogos formam pares resultando em um conjunto de quatro cromátides (tétrade). As cromátides mais próximas podem então “trocar” partes entre si. Sendo assim, os alelos antes ligados em uma cromátide, estarão separados, estando ligados aos genes de outra cromátide. Isto, em termos práticos, aumenta bastante a variabilidade genética.
O caso, meu querido, minha querida, é que, você provavelmente lembra que os cromossomos que formam o par de homólogos tiveram origem em cada um dos dois genitores. Sendo assim, durante a permuta, uma das cromátides do cromossomo que veio do pai se “mistura” com a que teve origem na mãe e assim formam uma nova combinação gênica aleatória. O crossing-over nem sempre acontece, sendo a formação de gametas parentais (iguais aos dos genitores) muito mais comuns.
Para entender melhor este processo, veja a imagem a seguir:
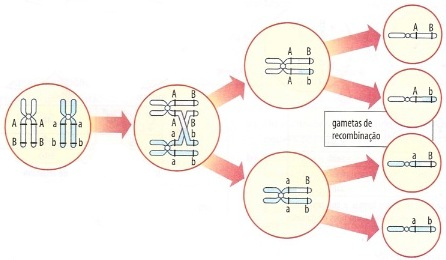
Vale relembrar: Quando dois pares de genes alelos estão situados muito próximos uns aos outros em um par de homólogos, é muito difícil que ocorra permutação entre eles. Diz-se, então, que há linkage total entre eles.
Caso os pares de genes analisados estão no mesmo cromossomo, porém mais distantes um do outro, há mais chance de que haja permutação, sendo assim, o linkage é parcial. Observe abaixo:
Se não houvesse recombinação nesses genes, a proporção de gametas formados por um duplo heterozigoto seria 50% AB e 50% ab. Quando há recombinação, observa-se na descendência uma pequena proporção de recombinantes, por exemplo:
40% - AB (parental)
40% - ab (parental)
10% - Ab (recombinante)
10% - aB (recombinante)
Quanto mais afastado um gene estiver do outro, maior será a taxa de recombinação.
Os gametas dos tipos AB a ab, cujo aparecimento não depende da ocorrência da permutação, são chamados gametas parentais, porque eles refletem a posição dos genes nas células. Os gametas dos tipos Ab e aB, que só aparecem caso aconteça a permutação, são chamados recombinantes.
3. Mapas gênicos
A partir da taxa de recombinação é possível construir o mapa gênico de um cromossomo. As unidades são medidas em unidades de recombinação (UR), morganídio ou centimorgan. Uma UR corresponde a 1% da taxa de recombinação.
Em drosófilas, encontraram as seguintes taxas de recombinação e mapearam os genes p, v e r:
4. RECONHECENDO UM CASO DE LINKAGE
Para reconhecermos um caso de linkage, é importante analisar os tipos de gameta formados por indivíduos heterozigotos e suas proporções. Imagine as seguintes situações:
Caso a) O indivíduo AaBb produz os seguintes gametas: ¼ AB; ¼ Ab; ¼ aB; ¼ ab.
Caso b) O indivíduo BbCc produz os seguintes gametas: ½ BC; ½ bc.
Caso c) O indivíduo CcDd produz os seguintes gametas: 40% CD, 10% Cd; 10% cD; 40% cd.
Veja que, no caso a, foram produzidos quatro gametas diferentes, todos na mesma proporção, indicando, assim, um caso claro de segregação independente. Já no caso b, formaram-se apenas dois gametas e ambos também com a mesma proporção. Sendo assim, esse caso representa um exemplo claro de linkage. Por fim, temos o caso c, que apresentou quatro gametas diferentes, porém em diferentes proporções. Quando isso acontece, dizemos que é um caso de linkage, porém com um diferencial de que é a ocorrência de uma permutação ou crossing-over.
5. Como calcular a taxa de crossing ou taxa de permutação
A taxa de crossing deve ser calculada a partir de dados de experimentos. Vamos usar como exemplo o cruzamento entre drosófilas (moscas de frutas).
Se cruzarmos uma drosófila híbrida, de corpo cinza e asa normal (NV/nv), com um macho de corpo preto e asa vestigial (nv/nv), vamos obter:
90 indivíduos com corpo preto e asa normal (nV/nv)
90 indivíduos com corpo cinza e asa vestigial (Nv/nv)
410 indivíduos com corpo preto e asa vestigial (nv/nv)
410 indivíduos com corpo cinza e asa normal (NV/nv)
Nesse caso, qual a taxa de crossing?

Retrocruzamento com genes ligados para cor do corpo e tamanho da asa da drosófila.
A primeira coisa a ser feita é identificar os indivíduos de recombinação, que são sempre aqueles resultantes de gametas que passaram por permutação. No nosso exemplo, esses gametas formaram os dois grupos de 90 indivíduos cada um. Assim temos um total de 180 recombinantes.
No retrocruzamento, a frequência de indivíduos recombinantes é igual à de gametas, portanto, basta transformar esse número em porcentagem. Temos um total de 1000 indivíduos (90+90+410+410), desse modo:
1000 – 100%
180 – x
x = 18%
Ou seja, a taxa de crossing nessas drosófilas é de 18%.
Vale ressaltar que se tivéssemos usado como exemplo um retrocruzamento em indivíduos diíbridos (com genes independentes), os quatro tipos de filhos teriam a mesma porcentagem: 25% ou ¼ para cada um. Dessa forma, nesses 1000 descendentes, teríamos 250 com cada característica.
========================================
O Sistema Genital
Mudanças no corpo
A descoberta do sexo acontece com a descoberta do corpo. Moças e rapazes costumam acompanhar atentamente as mudanças que ocorrem nos seus órgãos sexuais externos. Essas mudanças são provocadas pela ação de hormônios.
As características sexuais primárias, visíveis nos órgãos genitais, são determinadas geneticamente e estão presentes desde o nascimento, tanto no homem como na mulher.
O corpo masculino
As principais modificações visíveis no corpo masculino ao longo da adolescência estão descritas abaixo.
Os testículos (dentro do saco escrotal) crescem primeiro e, pouco tempo depois, o pênis. Na puberdade, os pelos surgem em diversos locais: no rosto, nas axilas, no peito e nas áreas próximas aos testículos. A voz também sofre mudanças.
Esse conjunto de características que se definem na puberdade, em consequência da ação hormonal, recebe o nome de características sexuais secundárias. Estas, porém, não obedecem a padrões rígidos. Adolescentes de mesma idade podem apresentar diferenças significativas em relação à estatura do corpo, quantidade de pelos, tamanho do pênis, timbre de voz etc. O grupo étnico a que pertence o indivíduo, a herança genética, hábitos alimentares, problemas de saúde, dentre outros fatores, são responsáveis por essas diferenças.
Assim, colegas de mesma idade que a sua podem ser mais altos ou mais baixos que você ou terem a voz mais ou menos grave que a sua, por exemplo. Isto não deve preocupá-lo. As pessoas são diferentes e apresentam ritmos desiguais de desenvolvimento do corpo. É importante gostar de você, aprendendo a cuidar e valorizar o seu próprio corpo.

Os rapazes possuem uma pequena quantidade de hormônios sexuais femininos, as garotas, uma pequena quantidade de hormônios sexuais masculinos. Na puberdade, às vezes, um pequeno desequilíbrio na quantidade desses hormônios pode provocar um ligeiro crescimento das mamas nos rapazes ou pelos em excesso nas garotas. Em geral, isso desaparece com o tempo, mas, se persistir, o mais aconselhável é procurar orientação médica.
Na região genital, encontramos o pênis e o saco escrotal.
Pênis e a Ejaculação – O pênis é um órgão de forma cilíndrica e constituído principalmente por tecido erétil, ou seja, que tem capacidade de se erguer. Com a excitação sexual, esse tecido e banhado e preenchido por maior quantidade de sangue, o que torna o pênis ereto e rígido. Na ponta do pênis, há a glande (a “cabeça”), que pode estar coberta pelo prepúcio.
Na glande, há o orifício da uretra, canal que no corpo masculino se comunica tanto com o sistema urinário quanto com o sistema reprodutor. O tamanho do pênis varia entre os homens e não tem relação biológica com fertilidade e nem com potência sexual.

***********************************************************
1. O que é ligação gênica?
Dois ou mais pares de genes alelos localizados em diferentes pares de cromossomos homólogos segregam-se independentemente. Portanto, esta é a condição de validade da segunda lei de Mendel.
Quando dois ou mais pares de genes alelos estão localizados em um mesmo par de cromossomos homólogos, eles não obedecem à lei da segregação independente. Afinal, durante a meiose irá haver uma tendência de que esses genes permaneçam unidos, quando o par de homólogos se separar, como mostra a figura abaixo.
Quando dois ou mais pares de genes alelos se localizam em um mesmo par de cromossomos, dizemos que eles apresentam ligação gênica (ou ligação fatorial). Os autores de língua inglesa dão a essa situação o nome de linkage. No entanto, há um fenômeno capaz de alterar essa tendência de união. É a permutação gênica (ou crossing-over), troca de fragmentos entre cromossomos homólogos, que pode acontecer na prófase da primeira divisão da meiose.
Quando dois pares de genes alelos estão situados de tal forma, em um par de homólogos, que não ocorre permutação entre eles, diz-se que há linkage total entre eles. Caso haja permutação, o linkage é parcial.
Em um caso de ligação gênica, não basta se conhecer o genótipo de um indivíduo. É necessário que se determine a posição relativa dos genes no par de homólogos. Por que isso é tão importante? Observe as duas situações mostradas a seguir:
Podemos notar que, embora as duas células possuam os mesmos genes, a sua posição, no par de cromossomos homólogos não é a mesma, o que determina a produção de tipos diferentes de gametas, na meiose.
Existem diversas formas de se indicar a posição dos genes no par de homólogos. As mais comuns são:
Uma outra forma de se indicar essa posição relativa dos genes é uma nomenclatura habitualmente usada pela química orgânica. O duplo-heterozigoto que tem os dois genes dominantes no mesmo cromossomo e os dois recessivos no outro (AB/ab) é chamado de heterozigoto "cis". O duplo-heterozigoto cujos genes dominantes estão em cromossomos diferentes do par de homólogos (Ab/aB) é o heterozigoto "trans".
2. Permutação ou crossing-over:
Teoricamente, os genes presentes em um mesmo cromossomo deveriam permanecer ligados uns aos outros, mesmo após a meiose. Porém, nem sempre isto ocorre. Muitas vezes é possível que os cromossomos troquem partes entre si durante a prófase da primeira fase da meiose.
Esse processo chamado de crossing-over ou permuta, ocorre quando os cromossomos homólogos formam pares resultando em um conjunto de quatro cromátides (tétrade). As cromátides mais próximas podem então “trocar” partes entre si. Sendo assim, os alelos antes ligados em uma cromátide, estarão separados, estando ligados aos genes de outra cromátide. Isto, em termos práticos, aumenta bastante a variabilidade genética.
O caso, meu querido, minha querida, é que, você provavelmente lembra que os cromossomos que formam o par de homólogos tiveram origem em cada um dos dois genitores. Sendo assim, durante a permuta, uma das cromátides do cromossomo que veio do pai se “mistura” com a que teve origem na mãe e assim formam uma nova combinação gênica aleatória. O crossing-over nem sempre acontece, sendo a formação de gametas parentais (iguais aos dos genitores) muito mais comuns.
Para entender melhor este processo, veja a imagem a seguir:
Vale relembrar: Quando dois pares de genes alelos estão situados muito próximos uns aos outros em um par de homólogos, é muito difícil que ocorra permutação entre eles. Diz-se, então, que há linkage total entre eles.
Caso os pares de genes analisados estão no mesmo cromossomo, porém mais distantes um do outro, há mais chance de que haja permutação, sendo assim, o linkage é parcial. Observe abaixo:
Se não houvesse recombinação nesses genes, a proporção de gametas formados por um duplo heterozigoto seria 50% AB e 50% ab. Quando há recombinação, observa-se na descendência uma pequena proporção de recombinantes, por exemplo:
40% - AB (parental)
40% - ab (parental)
10% - Ab (recombinante)
10% - aB (recombinante)
Quanto mais afastado um gene estiver do outro, maior será a taxa de recombinação.
Os gametas dos tipos AB a ab, cujo aparecimento não depende da ocorrência da permutação, são chamados gametas parentais, porque eles refletem a posição dos genes nas células. Os gametas dos tipos Ab e aB, que só aparecem caso aconteça a permutação, são chamados recombinantes.
3. Mapas gênicos
A partir da taxa de recombinação é possível construir o mapa gênico de um cromossomo. As unidades são medidas em unidades de recombinação (UR), morganídio ou centimorgan. Uma UR corresponde a 1% da taxa de recombinação.
Em drosófilas, encontraram as seguintes taxas de recombinação e mapearam os genes p, v e r:
| Genes | taxa de recombinação | Distância |
| p - v | 17% | 17 UR |
| p - r | 9% | 9 UR |
| r - v | 8% | 8 UR |
4. RECONHECENDO UM CASO DE LINKAGE
Para reconhecermos um caso de linkage, é importante analisar os tipos de gameta formados por indivíduos heterozigotos e suas proporções. Imagine as seguintes situações:
Caso a) O indivíduo AaBb produz os seguintes gametas: ¼ AB; ¼ Ab; ¼ aB; ¼ ab.
Caso b) O indivíduo BbCc produz os seguintes gametas: ½ BC; ½ bc.
Caso c) O indivíduo CcDd produz os seguintes gametas: 40% CD, 10% Cd; 10% cD; 40% cd.
Veja que, no caso a, foram produzidos quatro gametas diferentes, todos na mesma proporção, indicando, assim, um caso claro de segregação independente. Já no caso b, formaram-se apenas dois gametas e ambos também com a mesma proporção. Sendo assim, esse caso representa um exemplo claro de linkage. Por fim, temos o caso c, que apresentou quatro gametas diferentes, porém em diferentes proporções. Quando isso acontece, dizemos que é um caso de linkage, porém com um diferencial de que é a ocorrência de uma permutação ou crossing-over.
5. Como calcular a taxa de crossing ou taxa de permutação
A taxa de crossing deve ser calculada a partir de dados de experimentos. Vamos usar como exemplo o cruzamento entre drosófilas (moscas de frutas).
Se cruzarmos uma drosófila híbrida, de corpo cinza e asa normal (NV/nv), com um macho de corpo preto e asa vestigial (nv/nv), vamos obter:
90 indivíduos com corpo preto e asa normal (nV/nv)
90 indivíduos com corpo cinza e asa vestigial (Nv/nv)
410 indivíduos com corpo preto e asa vestigial (nv/nv)
410 indivíduos com corpo cinza e asa normal (NV/nv)
Nesse caso, qual a taxa de crossing?
Retrocruzamento com genes ligados para cor do corpo e tamanho da asa da drosófila.
A primeira coisa a ser feita é identificar os indivíduos de recombinação, que são sempre aqueles resultantes de gametas que passaram por permutação. No nosso exemplo, esses gametas formaram os dois grupos de 90 indivíduos cada um. Assim temos um total de 180 recombinantes.
No retrocruzamento, a frequência de indivíduos recombinantes é igual à de gametas, portanto, basta transformar esse número em porcentagem. Temos um total de 1000 indivíduos (90+90+410+410), desse modo:
1000 – 100%
180 – x
x = 18%
Ou seja, a taxa de crossing nessas drosófilas é de 18%.
Vale ressaltar que se tivéssemos usado como exemplo um retrocruzamento em indivíduos diíbridos (com genes independentes), os quatro tipos de filhos teriam a mesma porcentagem: 25% ou ¼ para cada um. Dessa forma, nesses 1000 descendentes, teríamos 250 com cada característica.
========================================
O Sistema Genital
Mudanças no corpo
A descoberta do sexo acontece com a descoberta do corpo. Moças e rapazes costumam acompanhar atentamente as mudanças que ocorrem nos seus órgãos sexuais externos. Essas mudanças são provocadas pela ação de hormônios.
As características sexuais primárias, visíveis nos órgãos genitais, são determinadas geneticamente e estão presentes desde o nascimento, tanto no homem como na mulher.
O corpo masculino
As principais modificações visíveis no corpo masculino ao longo da adolescência estão descritas abaixo.
Os testículos (dentro do saco escrotal) crescem primeiro e, pouco tempo depois, o pênis. Na puberdade, os pelos surgem em diversos locais: no rosto, nas axilas, no peito e nas áreas próximas aos testículos. A voz também sofre mudanças.
Esse conjunto de características que se definem na puberdade, em consequência da ação hormonal, recebe o nome de características sexuais secundárias. Estas, porém, não obedecem a padrões rígidos. Adolescentes de mesma idade podem apresentar diferenças significativas em relação à estatura do corpo, quantidade de pelos, tamanho do pênis, timbre de voz etc. O grupo étnico a que pertence o indivíduo, a herança genética, hábitos alimentares, problemas de saúde, dentre outros fatores, são responsáveis por essas diferenças.
Assim, colegas de mesma idade que a sua podem ser mais altos ou mais baixos que você ou terem a voz mais ou menos grave que a sua, por exemplo. Isto não deve preocupá-lo. As pessoas são diferentes e apresentam ritmos desiguais de desenvolvimento do corpo. É importante gostar de você, aprendendo a cuidar e valorizar o seu próprio corpo.
Os rapazes possuem uma pequena quantidade de hormônios sexuais femininos, as garotas, uma pequena quantidade de hormônios sexuais masculinos. Na puberdade, às vezes, um pequeno desequilíbrio na quantidade desses hormônios pode provocar um ligeiro crescimento das mamas nos rapazes ou pelos em excesso nas garotas. Em geral, isso desaparece com o tempo, mas, se persistir, o mais aconselhável é procurar orientação médica.
Na região genital, encontramos o pênis e o saco escrotal.
Pênis e a Ejaculação – O pênis é um órgão de forma cilíndrica e constituído principalmente por tecido erétil, ou seja, que tem capacidade de se erguer. Com a excitação sexual, esse tecido e banhado e preenchido por maior quantidade de sangue, o que torna o pênis ereto e rígido. Na ponta do pênis, há a glande (a “cabeça”), que pode estar coberta pelo prepúcio.
Na glande, há o orifício da uretra, canal que no corpo masculino se comunica tanto com o sistema urinário quanto com o sistema reprodutor. O tamanho do pênis varia entre os homens e não tem relação biológica com fertilidade e nem com potência sexual.
Quando o homem é estimulado, como ocorre numa relação sexual, culmina com o esperma sendo lançado para fora do corpo masculino sob a forma de jatos. Esse fenômeno chama-se ejaculação.
O esperma é ejaculado através da uretra, por onde a urina também é eliminada. Durante uma ejaculação normal são expelidos de 2 a 4 mililitros de esperma; cada mililitro contém aproximadamente 100 milhões de espermatozoides.
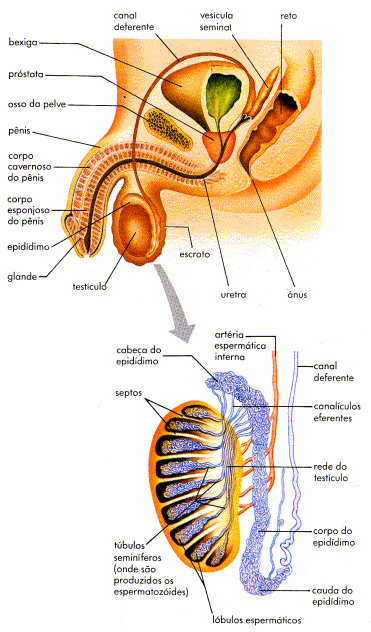
Saco escrotal
Os espermatozoides, gameta sexual masculino, são produzidos nos testículos. Os testículos ficam no saco escrotal, que tem aparência flácida e um pouco enrugada. É importante eles se localizarem fora do abdome, pois os espermatozoides são produzidos em uma temperatura mais baixa do que a do restante do corpo.
Nos dias frios ou durante um banho frio, o saco escrotal se encolhe, favorecendo o aquecimento dos testículos. O uso de cueca apertada pode causar infertilidade temporária, decorrente do aquecimento excessivo que provoca nos testículos.
Testículos
Os testículos são glândulas sexuais masculinas. São formadas por tubos finos e enovelados, chamados túbulos seminíferos. Diferentemente do que ocorre com as garotas, que já nascem com “estoque” de gametas (óvulos) “prontos” no corpo, é na puberdade, sob ação dos hormônios, que se inicia no corpo masculino a produção de gametas (os espermatozoides) nos testículos.
A produção de espermatozoides começa na puberdade, por volta dos 12 ou 13 anos de idade e vai até o fim da vida. Cada espermatozoide é formado basicamente de três partes: cabeça, colo e cauda com flagelo.
Os testículos produzem também o hormônio sexual masculino, chamado testosterona. O hormônio testosterona estimula o aparecimento das características sexuais secundárias masculinas: pelos no rosto e no restante do corpo, modificações na voz etc.
Epidídimos
Os espermatozoides que acabam de ser formados ficam armazenados no epidídimo, um outro enovelado de túbulos localizados sobre os testículos. Os epidídimos são dois órgãos formados por tubos enovelados, cada um localizado junto a um testículo. Reveja o esquema do sistema genital masculino e observe a localização dos epidídimos.
Os espermatozoides podem ficar armazenados nesses tubos por aproximadamente uma a três semanas, até que a maturação seja completada. Isso aumenta a sua mobilidade.
Os espermatozoides passam do epidídimo para um tubo com parede muscular chamado ducto deferente. De cada epidídimo parte um ducto deferente. Posteriormente e sob a bexiga urinária, cada ducto deferente se une ao canal da glândula seminal do mesmo lado e forma um tubo único, chamado ducto ejaculatório. Os ductos ejaculatórios lançam os espermatozoides num outro canal – a uretra. A uretra é um tubo que se inicia na bexiga urinária, percorre o interior do pênis e se abre no meio externo.
Glândulas Seminais e Próstata
As glândulas seminais são duas glândulas em forma de bolsa. Elas produzem um líquido denso que nutre os espermatozoides e aumenta a sua mobilidade.
A próstata é uma glândula produtora de um líquido de aspecto leitoso. Esse líquido é leitoso e neutraliza a acidez de restos de urina na uretra e, numa relação sexual, a acidez natural da vagina, protegendo assim os espermatozoides.
Em sua “viagem” até a uretra, os espermatozoides recebem os líquidos produzidos pelas glândulas seminais e pela próstata. Ao passar pela uretra, os espermatozoides recebem também um líquido lubrificante produzidos pelas glândulas bulbouretrais.
Ao conjunto formado pelos espermatozoides e os líquidos produzidos pelas glândulas seminais, pela próstata e pelas glândulas bulbouretrais dá-se o nome de esperma ou sêmen.
***********************************************************
Tecidos musculares
Os tecidos musculares são de origem mesodérmica e relacionam-se com a locomoção e outros movimentos do corpo, como a contração dos órgãos do tubo digestório, do coração e das artérias.
As células dos tecidos musculares são alongadas e recebem o nome de fibras musculares ou miócitos. Em seu citoplasma, são ricas em dois tipos de filamento protéico: os de actina e os de miosina, responsáveis pela grande capacidade de contração e distensão dessas células.
Quando um músculo é estimulado a se contrair, os filamentos de actina deslizam entre os filamentos de miosina. A célula diminui em tamanho, caracterizando a contração.
Tipos de tecido muscular
Há três tipos de tecido muscular: estriado esquelético, estriado cardíaco e liso. Cada um deles tem características próprias, adequadas ao papel que desempenham no organismo.

Tecido muscular estriado esquelético
O tecido muscular estriado esquelético constitui a maior parte da musculatura do corpo dos vertebrados, formando o que se chama popularmente de carne. Essa musculatura recobre totalmente o esqueleto e está presa aos ossos, daí ser chamada de esquelética. Esse tipo de tecido apresenta contração voluntária (que depende da vontade do indivíduo).
Um músculo esquelético é um pacote de longas fibras. Cada uma delas é uma célula dotada de muitos núcleos, chamado miócitos multinucleados. Um fibra muscular pode medir vários centímetros de comprimento, por 50 mm de espessura.

A célula muscular estriada apresenta, no seu citoplasma, pacotes de finíssimas fibras contráteis, as miofibrilas, dispostas longitudinalmente. Cada miofibrila corresponde a um conjunto de dois tipos principais de proteínas: as miosina, espessas, e as actinas, finas. Esses proteínas estão organizados de tal modo que originam bandas transversais, claras e escuras, características das células musculares estriadas, tanto as esqueléticas como as cardíacas.

Os filamentos de miosina formam bandas escuras, chamadas anisotrópicas (banda A), e os de actina, bandas claras, chamadas isotrópicas (banda I).
No centro de cada banda I aparece uma linha mais escura, chamada linha Z. O intervalo entre duas linhas Z consecutivas constitui um miômetro ou sarcômero e correspondem à unidade contrátil da célula muscular.
No centro de cada banda A existe uma faixa mais clara, chamada banda H, bem visível nas células musculares relaxadas e que vai desaparecendo à medida que a contração muscular ocorre.

Na contração muscular, os miofilamentos não diminuem de tamanho, mas os sarcômeros ficam mais curtos e toda a célula muscular se contrai.
O encurtamento dos sarcômeros ocorre em função do deslizamento dos miofilamentos finos sobre os grosso, havendo maior sobreposição entre eles: a banda I diminui de tamanho, pois os filamentos de actina deslizam sobre os de miosina, penetram na banda A e reduzem a largura da banda H.
A membrana plasmática da célula muscular estriada esquelética costuma ser chamada sarcolema (do grego, sarcos, carne).

Exercícios e o aumento da musculatura esquelética
Sabemos que exercícios físicos promovem o aumento da musculatura esquelética.
Mas o que aumenta: o número de células no músculo ou o volume das células já existentes?
A atividade física estimula as células musculares esqueléticas já existentes a produzirem novas miofibrilas, o que ocasiona aumento do volume da célula e conseqüentemente do músculo.

No indivíduo adulto, as células da musculatura esquelética não se dividem mais. No entanto, existem células especiais, chamadas satélites, que são mononucleadas e pequenas e se localizam no conjuntivo que envolve os miócitos. Em situações muito especiais, quando o músculo é submetido a exercícios intensos, essas células podem se multiplicar e algumas delas se fundir com as fibras musculares já existentes, contribuindo também para o aumento do músculo.
As células satélites são importantes nos processos de regeneração da musculatura esquelética quando ocorrem lesões.
Belo trabalho. Parabéns.
ResponderExcluir